

Arrierías 111
Autores:
Katherine Castaño Flórez1katherine.castanof@uqvitural.edu.co, https://orcid.org/0000-0002-7052-9685
Bióloga egresada de la Universidad del Quindío, Facultad de Ciencias Básicas y Tecnológicas, Armenia, Colombia.
Carlos Alberto Agudelo Henao agudelohenao@yahoo.com, https://orcid.org/0000-0002-1446-1639
M. Sc., Doctor en Ciencias Biología Universidad del Quindío, Facultad de Ciencias Básicas y Tecnologías, Programa de Biología, Armenia, Colombia.
Rafael Antonio Castaño Vélez ecoambientes3000@hotmail.com, https://orcid.org/0009-0004-5369-2503
Esp. Educación y gestión ambiental. Licenciado en Biología y Química, Universidad Santiago de Cali.
Artículo Publicado en Ciencia Latina Revista Científica Multidisciplinar, Ciudad de México, ISSN 2707-2207/ISSN 2707-2215, mayo -junio 2026. Volumen 10, Número 3.
DOI: https://doi.org/10.37811/cl_rcm.v10i3.24201
—

RESUMEN
El simbolismo en América ha estado presente a lo largo de la historia como expresión de lo cosmológico concreto, reflejando el pensamiento ancestral a través de animales representados en piezas arqueológicas. En Caicedonia -Valle del Cauca, este material constituye un indicador de la relación ecosistémica y cultural entre las comunidades prehispánicas y su entorno. Para tal efecto se desarrolló una investigación exploratoria, con el objetivo de identificar taxonómicamente las especies de animales representadas en piezas arqueológicas a través de mediciones de estructuras, guías y literatura especializada, lo que permitió establecer correspondencias con fauna actual, algunas en categoría de amenaza de acuerdo con listados nacionales. Se registraron trece piezas pertenecientes a los periodos Quimbaya Temprano I y Tardío II y III, con un total de cincuenta y ocho representaciones zoomorfas correspondientes a ranas, serpientes, aves y un lagarto.
Palabras clave: piezas arqueológicas, representaciones zoomorfas, taxonomía.
Taxonomy of the fauna represented in archaeological artifacts from the territory of Caicedonia, Valle del Cauca, Colombia
ABSTRACT
Symbolism in the Americas has been present throughout history as an expression of concrete cosmology, reflecting ancestral thought through animals depicted in archaeological artifacts. In Caicedonia, Valle del Cauca, this material constitutes an indicator of the ecosystemic and cultural relationship between pre-Hispanic communities and their environment. For this purpose, an exploratory study was conducted with the objective of taxonomically identifying the animal species represented in archaeological artifacts through structural measurements, guides, and specialized literature. This approach allowed the establishment of correspondences with present-day fauna, some of which fall under threat categories according to national listings. Thirteen artifacts from the Early Quimbaya I and Late Quimbaya II and
III periods were documented, comprising a total of fifty-eight zoomorphic representations corresponding to frogs, snakes, birds, and one lizard.
Keywords: archaeological artifacts, zoomorphic representations, taxonomy.
Artículo recibido 25 marzo 2026
Aceptado para publicación: 25 abril 2026
INTRODUCCIÓN
En América Latina, las sociedades prehispánicas se distinguían por elaborar diversos motivos de animales, representando la fauna y la flora de cada región (Redi, 2019; González, 1998). Estas expresiones forman parte del arte precolombino, que integra piezas arqueológicas, orfebrería, pintura, arquitectura y otros elementos de las artes indígenas, funcionando como vehículo de transmisión cultural y reflejando la cosmovisión de los pueblos originarios (Gamboa, 2014; González, 1998). La presente es una investigación interdisciplinaria, que abarca aspectos de etnozoología, taxonomía y arqueología, cada una de ellas proporciona los marcos de referencia que permiten reconocer ciertos objetos como “piezas arqueológicas”, sirviendo como evidencia material para reconstruir aspectos culturales y ambientales (Bovisio, 2013).
El estudio involucró índices de fidelidad, análisis de distancias euclidianas, el método de enlace de Ward que favorece la identificación de transiciones morfométricas entre los conjuntos analizados (Ward, 1963), análisis fenéticos, la validación de los agrupamientos jerárquicos mediante el método de multiscale bootstrap resampling. Todo implementado en los programas CONSEL y el paquete pvclust para R (Shimodaira & Hasegawa, 2001; Suzuki & Shimodaira, 2006).
En el ámbito regional, Duque (1970) realizó uno de los primeros estudios taxonómicos de la cultura Quimbaya a partir de material cerámico hallado en tumbas prehispánicas del Quindío y de piezas conservadas en el Museo del Oro de Bogotá, donde describió figuras de peces, anfibios, aves, reptiles y artrópodos. Posteriormente, Uribe-Villegas (1991) analizó la orfebrería Quimbaya Tardía, destacando técnicas y simbolismos que enriquecen la comprensión de las prácticas artísticas y culturales del Cauca Medio. En la misma línea, Legast (1993) examinó material precolombino Calima, identificando motivos de aves, mamíferos, peces y reptiles como parte de la interacción humano–fauna. En estudios más recientes, Uribe-Villegas (2016) analizó aves en la orfebrería del Cauca Medio, mientras que Gómez & Garrido (2017) abordaron la representación simbólica del jaguar y su vínculo con la conservación biológica. Finalmente, Castaño (2023) investigó la cerámica prehispánica de Caicedonia, hallando representaciones de anfibios, reptiles y aves en vasijas del periodo Quimbaya Tardío.
El interés surge del desconocimiento de la identificación taxonómica de la fauna representada en piezas arqueológicas del municipio de Caicedonia, Valle del Cauca, lo que dificulta la correcta identificación de los organismos, lo que puede derivar en errores al interpretar interacciones ecológicas y distribución geográfica; la diferenciación adecuada entre especies, generando confusión en la estimación de la biodiversidad y en la interpretación de datos zoológicos y genéticos; el avance en el conocimiento biológico de las especies, ya que la taxonomía constituye un paso previo e indispensable para estudios de ecología, genética y biogeografía; reconocer con certeza qué especies fueron plasmadas en las piezas, lo que produce un vacío en la comprensión del valor cultural y biológico de dichas representaciones; determinar con precisión la diversidad taxonómica representada, incluyendo la identificación a nivel de órdenes, familias y especies, lo cual resulta esencial para establecer patrones ecológicos y biogeográficos históricos; y articular la memoria arqueológica con el conocimiento científico de la biodiversidad local, limitando tanto la divulgación en los ámbitos educativo y comunitario. En consecuencia, la pregunta de investigación planteada fue: ¿Cuáles morfoespecies están representadas en las piezas arqueológicas del área de estudio y qué correspondencia tienen con la fauna actual de la región?
En este contexto, el objetivo del trabajo se orientó a la identificación taxonómica de la fauna representada en las piezas arqueológicas estudiadas, compararla con especies con posible distribución actual en la región.
METODOLOGÍA
El estudio se realizó en el municipio de Caicedonia, Valle del Cauca, Colombia, ubicado en la región Andina, que hace parte de la región sur fisiográfica del Quindío, específicamente en el flanco Occidental de la Cordillera Central, a una latitud de 4° 19′ 60» Norte y una longitud de 75° 49′ 42» Oeste (Municipios de Colombia, 2019).
Se visitó el Museo Arqueológico Prehispánico, perteneciente a la Casa de la Cultura Nacianceno Orozco Gallego (fig. 1), con el propósito de analizar las representaciones zoomorfas presentes en las piezas y compararlas con la fauna actual del municipio. Se seleccionaron las piezas que contenían representaciones de animales, a cada pieza se le asignó un código para facilitar su identificación y las morfoespecies fueron documentadas mediante fotografías.

Figura 1. Instalaciones del Museo Nacianceno Orozco, Caicedonia, Valle del Cauca.
Inicialmente se consultó literatura especializada para la elaboración de la lista de referencia de las especies actuales con posible distribución en el área de estudio (Castro-Herrera et al. 2007; Castro-Herrera & Vargas-Salinas 2008; Castro-Herrera & Bolívar-García 2010; Cardona-Botero et al. 2013; Acosta, 2017; Moreno Arias et al. 2023, entre otros) y la descripción taxonómica de cada grupo (Legast, 1998; Cardona-Botero et al. 2013; Agudelo-Valderrama et al. 2014; Vargas-Salinas et al. 2014; Fernández & Vanegas-Guerrero, 2017; Ríos-Soto el tal. 2018; Navarro-Salcedo et al. 2019; Atehortúa-Vallejo et al. 2020, entre otros). Seguidamente, se hicieron mediciones, utilizando un calibrador digital Steren ® (±0,01 mm), de las estructuras de cada individuo (Longitud rostro-cloaca (LCR), Ancho de la boca (AB), Longitud del antebrazo (LA), Longitud del fémur (LF), Longitud de la tibia-fíbula (LT) y se consideró Dato Faltante (DF), y finalmente, la identificación taxonómica.
Con el fin de cuantificar el grado de semejanza, se aplicó un análisis de distancias euclidianas en el software Infostat utilizando la distancia euclidiana y el método de enlace de Ward. Para individuos con datos faltantes (extremidades ausentes), se aplicó un procedimiento de estimación de datos faltantes, que permitió predecir valores ausentes basándose en la información disponible de las demás morfoespecies de ranas, evitando así eliminar individuos del análisis y manteniendo la comparabilidad entre todos los individuos. A partir de lo anterior se construyó el dendrograma para identificar las agrupaciones principales.
Para el análisis fenético de las morfoespecies de ranas, se llevó a cabo un análisis estadístico multivariado empleando el software R (versión 2025.09.1+401). La base de datos fue importada en formato .csv e incluyó una columna identificadora (Id) y un conjunto de variables correspondientes a las medidas obtenidas de cada morfoespecie. En primer lugar, se verificó la presencia de valores faltantes mediante un conteo general y, con el fin de evitar la exclusión de ejemplares, se aplicó un
proceso de imputación mediante el algoritmo missForest, basado en bosques aleatorios, que permite estimar valores faltantes de forma robusta sin alterar la estructura original del conjunto de datos. Posteriormente, los datos fueron estandarizados utilizando la función scale para garantizar que todas las variables tuvieran varianza unitaria y media cero, eliminando así el efecto de las diferencias de escala entre las medidas.
Con los datos estandarizados, se calculó una matriz de distancias euclidianas, la cual sirvió de base para la construcción de un dendrograma fenético mediante el método de agrupamiento jerárquico promedio no ponderado (UPGMA). Este análisis permitió representar las relaciones de similitud morfológica entre los ejemplares, mostrando la proximidad fenética en función de la distancia de las mediciones de las estructuras. El dendrograma fue posteriormente transformado en un objeto gráfico y personalizado visualmente mediante la librería dendextend, asignando grosores diferenciados a las ramas, colores a los grupos definidos y etiquetas legibles con los nombres originales de los ejemplares. Asimismo, se aplicó un análisis de validación por bootstrap con el paquete pvclust, que calculó valores de soporte Approximately Unbiased (AU) y Bootstrap Probability (BP) mediante 1000 repeticiones, brindando una estimación de la estabilidad de los agrupamientos obtenidos. Adicionalmente, se ejecutó un Análisis de Coordenadas Principales (PCoA o Classical Multidimensional Scaling), basado igualmente en la matriz de distancias euclidianas, con el fin de corroborar los patrones de similitud detectados y proporcionar una representación complementaria del espacio morfométrico. Complementariamente, se efectuó un Análisis de Componentes Principales (PCA) sobre los datos imputados y escalados, con el propósito de reducir la dimensionalidad del conjunto de variables y determinar los ejes que describen la mayor proporción de la variación en las mediciones de las estructuras. Los resultados del PCA se representaron mediante un biplot en el que se visualizaron simultáneamente los individuos (puntos) y las variables (vectores), permitiendo observar tendencias de agrupamiento y posibles gradientes de variación entre los cuencos de origen.
Para la identificación taxonómica se tomaron en cuenta la ausencia o presencia de patrones de líneas longitudinales. En cuanto a la identificación taxonómica de la morfoespecie de lagarto, se prestó atención a las proporciones corporales representadas, como las extremidades y la cola. Finalmente, para las morfoespecies de aves, debido a la dificultad de realizar mediciones, solo se tuvo en cuenta la disposición de las alas.
Para corroborar que las morfoespecies correspondieran al taxón asignado se consultaron libros y artículos de zoología (Salgado-Negret et al. 2016; Herrera, 2016; Cano, 2010) que describían detalladamente las partes corporales de los animales reales, incluyendo el número y disposición de extremidades, la forma de la cabeza y del torso, y otros rasgos característicos. Para cuantificar la correspondencia morfológica se aplicó la fórmula del Índice de Fidelidad de Friedman (1986): NF = (Np/N) *100 donde, Np= número de partes idénticas a los seres vivos con que se corresponden y N= número total de partes del individuo; dado que corresponde a una expresión aritmética básica, se adopta aquí como herramienta de análisis para las representaciones.
RESULTADOS
Tipos de piezas arqueológicas: Se registraron 13 piezas de cerámica con figuras zoomorfas, entre las que se cuentan ocho cuencos, tres ollas, una vasija y un volante de huso, correspondiendo dos de ellas al periodo temprano, tres al tardío I y cuatro al tardío II. Las piezas presentan un total de 58 representaciones de fauna, distribuidas en anfibios (anuros), reptiles (serpientes y lagarto) y aves. El análisis porcentual muestra que las morfoespecies de serpientes constituyen la mayoría con un 72,4 % del total, seguidas por las morfoespecies de anfibios (19 %), aves (6,9 %) y el lagarto (1,7 %).
Descripción morfológica de las morfoespecies de serpientes: Las representaciones de serpientes recorren el contorno del recipiente en un patrón de zigzag continuo (fig. 2 a) o en curvas en forma de “S”, pueden presentar su piel lisa o segmentada (fig. 2 b-e).

Figura 2. Detalle del aplique (a) con patrón de zigzag del cuenco en vista lateral. Cara número uno (b) y dos (c, d) de la olla con patrón en curvas en vista desde arriba (e).
Las morfoespecies O1S1, O3S1, V1S1 y C7S1 pueden pertenecer a tres géneros por el patrón segmentado a lo largo del cuerpo: Lampropeltis, Oxyrhopus y Micrurus.
En Caicedonia se ha registrado con frecuencia a Micrurus mipartitus, una serpiente venenosa de cuerpo delgado, cabeza poco diferenciada del cuello y anillos rojos o anaranjados en la cola (Ríos-Soto et al., 2018). En contraste, Oxyrhopus petolarius es una falsa coral que imita a la coral verdadera mediante mimetismo, diferenciándose por tener la cabeza más definida y bandas negras y claras alternadas en el cuerpo (Acosta-Ortiz et al., 2021). Por último, en la región posiblemente se encuentra Lampropeltis triangulum, reconocida por su patrón repetitivo de anillos amarillos, negros y rojos desde el cuello (Williams, 1994). Su tamaño varía según la región, siendo menor en el sureste de EE. UU. y mayor en México y Centroamérica (Williams, 1994). La forma de la cabeza también puede variar, desde poco diferenciada hasta muy marcada respecto al cuerpo (Williams, 1994).
Para las morfoespecies O2S1 y O2S2 se sugiere una posible asociación con los géneros Clelia y Trilepida, respectivamente. En el contexto del municipio de Caicedonia, las especies que podrían corresponder a Clelia clelia y Trilepida joshuai. Esta propuesta se basa en que, estas serpientes presentan patrones de coloración distintivos, como líneas, franjas u otras marcas, tanto C. clelia como T. joshuai se caracterizan por una coloración más uniforme a lo largo del cuerpo.
Descripción morfológica de la morfoespecie de lagarto: Se encuentra adherido al borde del cuenco en una postura que sugiere movimiento ascendente, con las extremidades posteriores e inferiores en contacto con la superficie y la cola extendida a lo largo del borde (fig. 3). La disposición de su cuerpo transmite la idea de escalada o desplazamiento activo, en contraste con las representaciones en posición de reposo.

Figura 3. Cara número uno (a, b) del cuenco con aplique zoomorfo de lagarto en vista desde arriba (c).
La morfoespecie C8L1 podría corresponder al género Anolis, debido a sus extremidades delgadas y alargadas, rasgo distintivo frente a otros géneros presentes en Caicedonia. Especies como Anolis heterodermus presentan hábitos arborícolas y habitan matorrales y bosques andinos (Ríos-Orjuela et al., 2020), mientras que Anolis antonii comparte características como cuerpo pequeño, extremidades largas y hocico corto (Catálogo de la Biodiversidad, s.f.). Su amplia distribución y cercanía a zonas habitadas sugieren que pudieron ser observadas con frecuencia por poblaciones prehispánicas y representadas en piezas arqueológicas. Esta interpretación fue respaldada por Vargas-Salinas, F. (com. pers., 05 de marzo de 2025).
Descripción morfológica de las morfoespecies de anuros: Se encuentran adheridos al borde de cada cuenco, representando a los individuos en posición de reposo. Algunos muestran cabeza pronunciada y ojos abultados; las extremidades delanteras se extienden hacia los lados, mientras que las traseras están dobladas y ligeramente separadas del dorso (fig. 4).

Figura 4. Cara número uno a detalle (a, b) y dos (c,d) del cuenco con apliques de anuros en vista desde arriba (e).
Análisis Integrado de Distancias Euclidianas y Fenético (fig. 5)
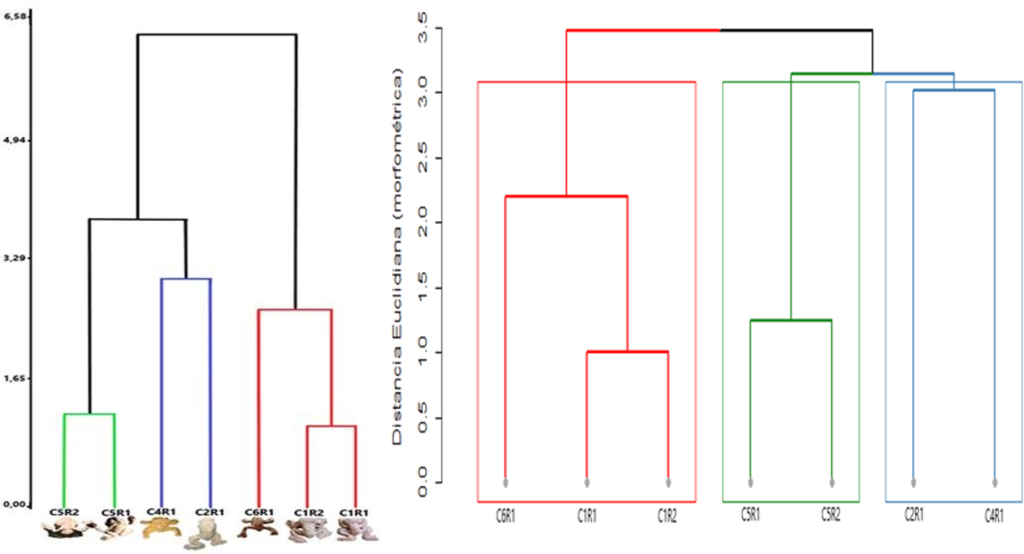
Figura 5. Dendrogramas obtenidos del análisis de las morfoespecies de ranas de los cuencos: a) Dendrograma de la izquierda con tres agrupaciones de morfoespecies de ranas obtenidas a partir de distancias euclidianas, que incluye una imagen de los individuos representados en los cuencos. b) Dendrograma fenético de la derecha basado en distancias euclidianas estandarizadas entre los ejemplares analizados.
El dendrograma basado en distancias euclidianas mostró tres agrupaciones principales entre los ejemplares analizados, donde las muestras con proporciones corporales similares formaron conjuntos compactos y homogéneos (Lele & Richtsmeier, 1991). Este patrón indica que las diferencias observadas responden a variaciones morfológicas reales y no aleatorias, coincidiendo con estudios previos sobre análisis de forma y tamaño (Forina et al., 2002; Murtagh & Contreras, 2012). Además, estas agrupaciones se mantuvieron de forma consistente en el dendrograma obtenido mediante el método UPGMA, conservando la jerarquía y los niveles relativos de similitud entre los ejemplares, lo que evidencia estabilidad en las relaciones fenéticas independientemente de la representación utilizada.
Validación del agrupamiento: Agrupamiento jerárquico (fig. 6)
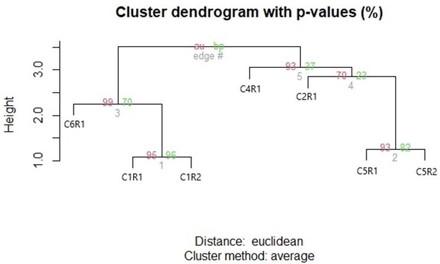
Figura 6. Agrupamiento jerárquico de siete morfoespecies de ranas, validado mediante multiscale bootstrap resampling (n = 1000) mostrando valores de soporte AU y BP.
En el dendrograma se evidenciaron tres clados principales que coinciden plenamente. El primer clado, conformado por C5R2 y C5R1 en color verde, se ubica muy próximo al valor cero de distancia, con AU
= 93% y BP = 82%, lo que refleja un soporte estadístico moderadamente alto y una alta similitud en la estructura corporal entre ambos ejemplares. Estos individuos presentan proporciones casi idénticas en las longitudes corporales y de las extremidades (fig. 7), lo que sugiere que pertenecen a la misma morfoespecie o que comparten un conjunto de rasgos muy específicos, como la morfología de la cabeza y la LRC.
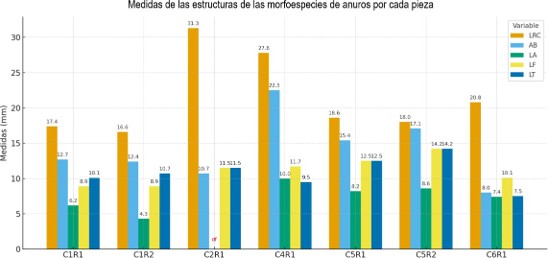
Figura 7. Medidas de las estructuras de las morfoespecies de anuros por cada pieza.
El segundo clado, representado por C4R1 y C2R1 en color azul, se agrupa a una distancia mayor con AU = 93% y BP = 37%, lo que indica un soporte moderadamente alto bajo el criterio AU. Esta variación refleja una mayor heterogeneidad en la estructura entre ambos ejemplares, los cuales presentan las mayores longitudes rostro-cloaca del conjunto, junto con una mayor proporción corporal. Aunque C2R1 es ligeramente más grande que C4R1, ambos comparten proporciones semejantes en la longitud total (fig. 7), lo que explica su agrupamiento. La menor consistencia estadística (BP) podría estar dada por la falta de información completa en alguna medida, como la extremidad de C2R1.
El tercer grupo, identificado por líneas rojas y conformado por C6R1, C1R2 y C1R1, muestra una subdivisión interna claramente soportada estadísticamente. Los ejemplares C1R1 y C1R2 presentan AU
= 95% y BP = 96%, lo que indica una coincidencia alta en sus proporciones corporales, sugiriendo una marcada homogeneidad. A este subgrupo se une C6R1 con valores AU = 99% y BP = 70%, lo que refleja una afinidad en las medidas estructurales alta, pero con ligeras diferencias. Aunque comparte rasgos generales de tamaño corporal reducido con las morfoespecies C1R1 y C1R2, C6R1 presenta diferencias sutiles, principalmente en la conformación de la cabeza y las proporciones de las extremidades (fig. 7), lo que explica su unión a una distancia ligeramente mayor.
Análisis de Coordenadas Principales (PCoA) Fenético (fig. 8)
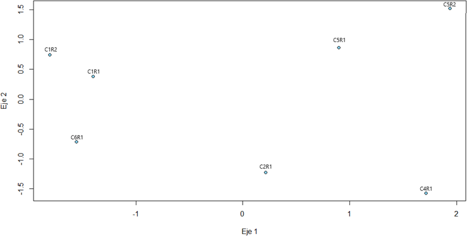
El análisis de coordenadas principales (PCoA) (fig. 8) permitió representar las relaciones fenéticas entre las morfoespecies de anuros a partir de la matriz de distancias euclidianas, calculadas con base en las variables morfométricas estandarizadas. Este método busca proyectar en dos o más ejes las posiciones
relativas de los individuos según sus distancias mutuas, de modo que aquellos ejemplares morfológicamente similares se ubiquen próximos en el espacio bidimensional (Gower, 1966; Legendre & Legendre, 2012). En la gráfica resultante (fig. 8), los dos ejes describen conjuntamente la mayor parte de la variabilidad fenética del conjunto de datos, reflejando de manera espacial la similitud o disimilitud entre los ejemplares analizados.
En el Eje 1, que representa el principal gradiente de disimilitud en las mediciones de las estructuras, se observa una clara separación entre los grupos ubicados hacia los valores negativos (C1R1, C1R2 y C6R1) y aquellos localizados hacia los valores positivos (C2R1, C4R1, C5R1 y C5R2). Esta distribución coincide con los agrupamientos obtenidos previamente en los dendrogramas, indicando que las relaciones observadas son consistentes independientemente del método empleado.
El primer grupo, conformado por C1R1, C1R2 y C6R1, se sitúa en el cuadrante izquierdo del gráfico, evidenciando una alta similitud fenética entre los dos primeros (C1R1 y C1R2), los cuales aparecen muy próximos entre sí, mientras que C6R1 se ubica ligeramente desplazado hacia valores negativos del eje
2. Esto sugiere que, aunque comparte rasgos en las estructuras generales con los dos primeros, presenta ciertas diferencias proporcionales en la conformación corporal.
El segundo grupo, formado por C5R1 y C5R2, se localiza en el extremo derecho superior del plano, con una distancia mínima entre ambos ejemplares. Esta proximidad indica una casi total correspondencia en sus proporciones corporales, lo que refuerza la idea sobre pertenecer a una misma morfoespecie.
Por su parte, los ejemplares C2R1 y C4R1 se ubican en el cuadrante derecho inferior, a una distancia intermedia del grupo C5R1-C5R2. Aunque ambos muestran ciertas similitudes en tamaño corporal general, la separación espacial sugiere diferencias marcadas en otras de sus estructuras, lo que respalda su posible asignación a morfoespecies distintas.
Análisis del PCA Fenético (fig. 9)
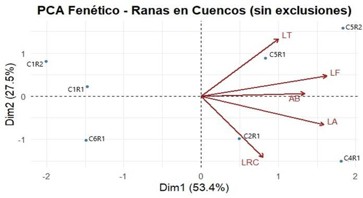
Figura 9. Análisis de Componentes Principales (PCA)
El análisis de Componentes Principales (PCA) fenético (fig. 9) permitió describir la variación en las estructuras entre los ejemplares de ranas de los distintos cuencos, considerando las variables de LRC, AB, LA, LF y LT. Este método multivariado reduce la dimensionalidad del conjunto de datos al transformar las variables originales en un nuevo sistema de coordenadas (componentes principales) que maximizan la varianza explicada y permiten identificar patrones subyacentes de similitud o diferenciación morfológica entre individuos (Jolliffe & Cadima, 2016; Legendre & Legendre, 2012). El primer componente (Dim1) explicó el 53,4 % de la variación total, mientras que el segundo componente (Dim2) explicó el 27,5 %, acumulando un 80,9 % de la variación total (fig. 9).
En el biplot obtenido, las flechas que representan las variables en la estructura se orientaron principalmente hacia el lado derecho de la Dim1, lo que indica una correlación positiva entre todas las medidas corporales, reflejando que los individuos con mayores dimensiones en una característica tienden a presentar incrementos proporcionales en las demás. Esta tendencia sugiere que Dim1 representa un gradiente de tamaño corporal total, donde los ejemplares se distribuyen de acuerdo con su magnitud general.
Las variables LRC y LA se proyectaron hacia la parte inferior derecha del gráfico, indicando que contribuyen negativamente la Dim2 y que su mayor variación se asocia con individuos de mayor longitud corporal, como los ejemplares C2R1 y C4R1, que en la tabla de las mediciones de las estructuras registran las mayores medidas de longitud rostro cloaca. Por el contrario, las variables LT, LF y AB, ubicadas en la parte superior derecha, presentan contribuciones positivas a la Dim2, vinculándose con proporciones relativamente mayores en las extremidades y la cabeza. Estas variables
se relacionan con los ejemplares C5R1 y C5R2, situados en el cuadrante superior derecho, que muestran
valores elevados de LF y LT, indicando extremidades más desarrolladas en relación con su tamaño corporal total.
Los ejemplares C1R1, C1R2 y C6R1 se distribuyeron hacia la parte izquierda del gráfico, asociándose con los valores más bajos de la Dim1, lo que refleja dimensiones corporales menores en comparación con los demás ejemplares. Este grupo se caracteriza por longitudes de rostro-cloaca más reducidas, así como menores valores en las medidas de extremidades.
Descripción morfológica de cada morfoespecie de rana
En cuanto a la descripción morfológica individual, se tiene que C5R2 y C5R1 tienen una medida corporal (LRC) entre 18 y 18,6 mm, lo que podría relacionarlos con ranas del género Pristimantis, cuyas especies registran tamaños entre 15 y 71 mm de longitud rostro-cloaca, según especie y sexo (Atehortúa-Vallejo et al., 2020). Además, el hocico redondeado y las extremidades extendidas con dedos marcados mediante incisiones simples del aplique podrían representar rasgos típicos del género, haciendo alusión a los discos expandidos en los dedos de la rana real (Atehortúa-Vallejo et al., 2020; Navarro-Salcedo et al., 2019; Fernández & Vanegas-Guerrero, 2017).
Seguido de este grupo, se encuentra C4R1 y C2R1 donde se sugiere que C2R1 puede ser asignada a la familia Hylidae, por sus medidas corporales que alcanzan hasta los 31,3 milímetros y por sus extremidades robustas y largas (Agudelo-Valderrama et al. 2014; Ortiz et al. 2022), mientras que C4R1, de extremidades más delgadas y esbelta morfología, se aproxima al patrón típico de los Centrolénidos (Cochran, 1968 citado en Legast, 1998).
Dentro de la familia Hylidae, en Caicedonia podrían estar presentes los géneros Hyloscirtus y Dendropsophus (Cadavid et al., 2005; Castro-Herrera & Vargas-Salinas, 2008; Castro-Herrera & Bolívar-García, 2010; Cardona-Botero et al., 2013). No obstante, las especies de Hyloscirtus que pueden estar registradas para la zona tienen dimensiones que superan las del ejemplar, por lo que su correspondencia es poco probable (Yánez-Muñoz et al., 2021; Ortiz et al., 2022).
En cambio, Dendropsophus columbianus se ajusta mejor a las dimensiones de la pieza, ya que sus machos miden entre 25,8 y 29,3 mm y las hembras hasta 35,4 mm de LRC. Además, presenta hocico corto y redondeado, así como extremidades posteriores largas (Agudelo-Valderrama et al., 2014). Según Vargas-Salinas (com. pers., 5 de marzo de 2025), esta especie pudo haber sido abundante cerca de antiguos asentamientos indígenas, facilitando su observación frecuente.
El ejemplar C4R1 podría relacionarse con la familia Centrolenidae, presente en Caicedonia con géneros como Centrolene, Nymphargus e Hyalinobatrachium (Castro-Herrera & Vargas-Salinas, 2008; Castro-Herrera & Bolívar-García, 2010; Cardona-Botero et al., 2013). Sin embargo, Hyalinobatrachium tatayoi resulta demasiado pequeño para corresponder a la pieza, y especies de Centrolene, aunque de tamaño similar, presentan hocico redondeado. Por ende, la mejor candidata sería Nymphargus griffithsi, registrada en zonas cercanas como La Tebaida, Armenia y Valle del Cauca, lo que hace plausible su presencia en Caicedonia (Acosta, 2017; Cadavid et al., 2005; Guayasamín et al., 2023). Sus medidas corporales coinciden con las del ejemplar (oscila entre los 20 y 27,8 mm de LRC) y posee un hocico truncado, rasgo también observado en la figura zoomorfa (Guayasamín et al., 2023).
Posteriormente, tenemos a C6R1, con cabeza poco diferenciada del torso, extremidades delgadas y largas, y una LRC de 20,8 mm, podría estar inspirado en la familia Dendrobatidae, especialmente en los géneros Hyloxalus, Colostethus y Andinobates. Sin embargo, las especies de Colostethus que pueden estar presentes en la región suelen presentar cabezas más amplias (Guevara-Molina et al. 2017), y las de Hyloxalus extremidades más cortas y robustas (Castro-Herrera & Bolívar-García, 2010). En contraste, el aplique coincide mejor con Andinobates bombetes, caracterizada por extremidades delgadas y una cabeza ligeramente más angosta que el cuerpo, siendo un caracter diagnóstico de la especie (Vargas-Salinas et al., 2014; Vargas-Salinas et al., 2020).
Por otra parte, el análisis estadístico agrupó los ejemplares C1R1 y C1R2 con A. bombetes, pero al comparar sus dimensiones y morfología con especies registradas en Caicedonia surgieron discrepancias. Se descartó su relación con familias como Centrolenidae, Hylidae, Strabomantidae, Leptodactylidae y Bufonidae, así como con A. bombetes y Hyloxalus, debido a diferencias de tamaño y rasgos corporales (Legast, 1993; Legast 1998; Barona-Cortés et al., 2007; Heyer et al., 2006; Castro-Herrera & Bolívar-García, 2010; Vargas-Salinas et al., 2014; Agudelo-Valderrama et al., 2014; Ortiz et al., 2022). En consecuencia, no fue posible asociarlos con una especie concreta de la región. Sin embargo, podrían corresponder a una especie localmente extinta desde época prehispánica o a una aún no descrita en Caicedonia.
Finalmente, se encuentran las morfoespecies C3R1 y C3R2, cuya identificación taxonómica resultó
compleja debido a que en la representación plasmada en la vasija solo se conserva la mitad del cuerpo del anfibio. Sin embargo, a partir de las características visibles como extremidades posteriores robustas y un torso aparentemente grueso o fornido, es posible inferir que puede tratarse de un ejemplar con un cuerpo compacto y macizo de la familia Bufonidae, como lo es la especie Rhinella marina con posible distribución en Caicedonia (Legast, 1993; Legast, 1998). A pesar de esta hipótesis preliminar, la falta de información anatómica completa impide asignar con certeza el ejemplar a un género o especie específica.
Descripción morfológica de las morfoespecies de aves: Se disponen de manera que sugieren una postura de vuelo o desplazamiento aéreo, posiblemente capturando el momento en que el ave se encuentra en pleno movimiento. Sin embargo, la estilización de sus formas es tal que impide establecer con certeza una relación directa con un género o especie (fig. 10).

Figura 10. Cara número uno (a) y dos (b) del volante de huso con representaciones de aves en vista desde arriba (c).
Podría tratarse de un ave rapaz, dadas las proporciones corporales, como la masa alar y la longitud de los huesos del ala, que mantienen una relación alométrica con el tamaño corporal y generan rasgos distintivos frente a otras aves, además de la posible sugerencia de alas extendidas y alargadas (Márquez et al., 2005; Bribiesca-Contreras et al. 2019). Asimismo, estas aves han sido símbolos centrales en el surgimiento de diversas culturas, identidades colectivas, movimientos ideológicos y expresiones de poder y organización social (Márquez et al., 2005). No obstante, también es plausible que corresponda a un ave común del entorno cotidiano, interpretada con libertad estética por parte del alfarero.
En total, de las 32 especies de anuros con posible distribución en Caicedonia, cuatro se encuentran representadas en las piezas arqueológicas y corresponden a especies aún presentes en la región donde, según la UICN Nymphargus griffithsi está en preocupación menor y Andinobates bombetes se encuentra
en estado vulnerable. En cuanto a las serpientes, de las 14 especies esperadas, cinco aparecen plasmadas en las representaciones y también se mantienen vigentes en la actualidad. Finalmente, dentro de las 15 especies de lagartos con posible distribución en el municipio, uno está representado en la pieza y corresponde igualmente a una morfoespecie vigente.
Fidelidad de las morfoespecies: Los resultados muestran que las serpientes y los anuros fueron las morfoespecies más representadas en las piezas arqueológicas de Caicedonia. En el caso de los anuros, el 66,7 % de las representaciones alcanzó un 100 % de fidelidad anatómica, lo que evidencia una observación detallada y una intención tanto naturalista como simbólica, similar a la registrada en la cerámica Calima y Yotoco (Legast, 1993). Su importancia cultural se relaciona con su simbolismo en el mundo andino, donde los anuros eran asociados con la Pachamama, la fertilidad, el agua y la renovación de la vida (Ochante-Sauñe, 2015). Además, su metamorfosis representaba transformación y renacimiento (Uribe-Villegas, 1991), mientras que en la Amazonía su canto era interpretado como señal de lluvias y abundancia agrícola (Ochante-Sauñe, 2015).
Las morfoespecies de serpientes mostraron un bajo nivel de fidelidad anatómica, ya que solo el 16,7 % de las figuras reprodujeron con exactitud los rasgos reales del animal. Esto sugiere que los Quimbayas priorizaron el simbolismo sobre la representación anatómica, al igual que otras culturas como la Calima, donde las serpientes se asociaban con figuras antropomórficas y otros animales (Legast, 1993).
En distintas culturas indígenas colombianas, las serpientes tuvieron significados relacionados con la creación, la fertilidad, la transformación y el equilibrio del mundo. Por ejemplo, para los Pijao la Serpiente Molá representaba la fuerza creadora (Tocarema, 2018), mientras que en las mitologías kogui y muisca simbolizaban la muerte, el renacimiento, el agua y la continuidad de la vida (Legast, 2000). La morfoespecie de lagarto alcanzó un índice de fidelidad del 100%, indicando que las estructuras corporales de la pieza coinciden completamente con la anatomía real. Además, este tipo de representación también habría estado presente en la cultura Calima, lo que sugiere una posible continuidad cultural (Legast, 1993). En las culturas Muisca y Quimbaya, el lagarto se relacionaba con el agua, la fertilidad, la abundancia y la muerte, especialmente en contextos funerarios y rituales (Chaves, 1981).
Por su parte, las morfoespecies de aves obtuvieron un índice de fidelidad del 60%, mostrando una correspondencia parcial con la morfología real. En el mundo prehispánico colombiano, las aves tuvieron un importante valor simbólico dentro del chamanismo, ya que su capacidad de volar se asociaba con el trance espiritual y el acceso a planos sagrados (Cuenca, 2011).
CONCLUSIONES
La identificación taxonómica de la fauna representada en las piezas arqueológicas recolectadas en el municipio de Caicedonia permitió reconocer las especies que tuvieron relevancia para las comunidades prehispánicas, reflejando un conocimiento detallado del entorno natural. Además, se comprobó que varias de estas especies posiblemente cuentan con representantes actuales en la región, lo que evidencia una continuidad tanto ecológica como cultural entre el pasado y el presente. A partir de estos hallazgos, se implementaron diversas estrategias de divulgación como conferencias, charlas en medios radiales, participación en eventos científicos y arqueológicos, y la elaboración de una cartilla pedagógica, que facilitaron la apropiación social del conocimiento y el fortalecimiento del vínculo entre la comunidad y su patrimonio.
Agradecimientos
A la Universidad del Quindío por su respaldo académico, a la Alcaldía de Caicedonia por facilitar el ingreso y el acceso a las piezas arqueológicas del Museo de la Casa de la Cultura Nacianceno Orozco, a Fernando Vargas por su apoyo científico, al Grupo de Estadística de la Universidad del Quindío y a la comunidad de Caicedonia por su participación.
Conflicto de intereses: Los autores declaramos que no tenemos conflictos de interés.
REFERENCIAS BIBLIOGRÁFICAS
Acosta, A. R. (2017). Nymphargus griffithsi. En Batrachia: Lista de los Anfibios de Colombia. https://www.batrachia.com/orden-anura/centrolenidae-82-spp/nymphargus-griffithsi/
Acosta-Ortiz, J. M., Bobadilla-Molina, J. S., Sierra-Rueda, A. S., & Astwood-Romero, J. A. (2021). Oxyrhopus petolarius (Linnaeus, 1758) – falsa coral. Catálogo de Anfibios y Reptiles de Colombia, 7(1), 39–50.
Agudelo-Valderrama, O. L., Bolívar-G, W., & Hernández-Medina, C. A. (2014). Dendropsophus columbianus. Catálogo de Anfibios y Reptiles de Colombia, 2(1), 40–44.
Alvarado, R. (1966). Sistemática, taxonomía, clasificación y nomenclatura. COL-PA, 9, 3–8. Atehortúa-Vallejo, M. A., Montoya-Marín, M., López-Molina, K. J., & Duarte-Marín, S. (2020).
Pristimantis erythropleura: Rana de ingle roja. Asociación Colombiana de Herpetología –
ACH. Catálogo de Anfibios y Reptiles de Colombia, 6(1), 43–50.
Barona-Cortés, E., & Mendoza-Henao, A. M. (2007). Hyalinobatrachium tatayoi: Rana de cristal de Tatayo. Catálogo de Anfibios y Reptiles de Colombia, 8(1), 43–54.
Bovisio, M. A. (2013). El dilema de las definiciones ontologizantes: Obras de arte, artefactos etnográficos, piezas arqueológicas. Caiana: Revista de Historia del Arte y Cultura Visual, (3), 1–10.
Bribiesca-Contreras, F., Parslew, B., & Sellers, W. I. (2019). A quantitative and comparative analysis of the muscle architecture of the forelimb myology of diurnal birds of prey (order Accipitriformes and Falconiformes). The Anatomical Record, 302(10), 1808–1823. https://anatomypubs.onlinelibrary.wiley.com/doi/10.1002/ar.24195
Brower, J. C. (2008). Euclidean distances and singular value decomposition: Useful tools for geometric morphometrics in biology and paleontology. In P. Frankl & N. Stoffer (Eds.), Progress in geomathematics (pp. 393–415). Springer. https://doi.org/10.1007/978-3-540-69496-0_20
Burbano-Vallejo, E. L., & Moreno, E. (2018). Análisis de conglomerados del norte del Valle del Cauca: Caso estudio Cartago, Zarzal y La Unión. Ingeniería Industrial, 39(1), 78–91. http://scielo.sld.cu/scielo.php?pid=S1815-59362018000100009&script=sci_arttext
Cardona-Botero, V. E., Viáfara-Vega, R. A., Valencia-Zuleta, A., Echeverry-Bocanegra, A., Hernández-Córdoba, O. D., Jaramillo-Martínez, A. F., … & Castro-Herrera, F. (2013). Diversidad de la herpetofauna en el Valle del Cauca (Colombia): Un enfoque basado en la distribución por ecorregiones, altura y zonas de vida. Biota Colombiana, 14(2).
Castaño, R. A. (2023). ¿Qué tipos de animales sagrados existieron en Caicedonia? Arrierías. https://www.arrierias.com/que-tipos-de-animales-sagrados-existieron-en-caicedonia-por-rafa-davidzen/
Castro, A. B. S. (2022). La ¿limitada? capacidad de generalización de los hallazgos de la investigación cualitativa. NURE Investigación: Revista Científica de Enfermería, (120), 1–3. https://dialnet.unirioja.es/servlet/articulo?codigo=8584897
Castro-Herrera, F., & Bolívar-García, W. (2010). Libro rojo de los anfibios del Valle del Cauca.
Corporación Autónoma Regional del Valle del Cauca (CVC).
Castro-Herrera, F., Bolívar-García, W., & Herrera-Montes, M. I. (2007). Guía de los anfibios y reptiles del bosque de Yotoco, Valle del Cauca, Colombia. Grupo de Investigación Laboratorio de Herpetología, Universidad del Valle.
Castro-Herrera, F., & Vargas-Salinas, F. (2008). Anfibios y reptiles en el departamento del Valle del Cauca, Colombia. Biota Colombiana, 9(2).
Catálogo de la Biodiversidad. (s.f.). Anolis antonii. https://catalogo.biodiversidad.co/file/56cfbd973c16479905cba7fe/
Chaves, Á. (1981). Los animales mágicos en las urnas de Tierradentro. Museo de Artes y Tradiciones Populares.
Corporación Autónoma Regional del Quindío. (2011). Plan de manejo subcuenca río Quindío.
Corporación Autónoma Regional del Valle del Cauca (CVC) & Funagua. (2010). Ecosistemas del Valle del Cauca. Informe final convenio 256/2009. Santiago de Cali.
Cuenca, J. V. R. (2011). Cosmovisión, chamanismo y ritualidad en el mundo prehispánico de Colombia: Esplendor, ocaso y renacimiento. Maguaré, 25(2), 145–195.
Dagnino, J. (2014). Datos faltantes (missing values). Revista Chilena de Anestesia, 43, 332–334. https://revistachilenadeanestesia.cl/PII/revchilanestv43n04.13.pdf
Díaz-Ricaurte, J. C., Serrano, F., & Ferreto Fiorillo, B. (2018). Clelia clelia (Daudin, 1803): Cazadora negra, chonta, culebra minadora, lisa, tiznada. Catálogo de Anfibios y Reptiles de Colombia, 4(2), 23–31.
Duque, L. (1970). Los Quimbayas. Edinalco Ltda.
Efron,B. (1979) Bootstrap methods: another look at the jackknife. Ann. Stat., 7, 1–26.
Efron, B., & Tibshirani, R. J. (1994). An introduction to the bootstrap. Chapman and Hall/CRC.
Felsenstein, J. (1985) Confidence limits on phylogenies: an approach using the bootstrap. Evolution, 39, 783–791.
Fernández, J. D., & Vanegas-Guerrero, J. (2017). Pristimantis piceus. Anfibios y Reptiles, 12.
Fernández, M. S., Brusa, F., Damborenea, M. C., Dellapé, P. M., & Gallardo, F. E. (2013). Introducción a la taxonomía: Manual de ejercitaciones (1.ª ed.). Editorial de la Universidad de La Plata.
Forina, M., Armanino, C., & Raggio, V. (2002). Clustering with dendrograms on interpretation variables. Analytica Chimica Acta, 454(1), 13–19. https://doi.org/10.1016/S0003-2670(01)01517-3
Friedman, J., Yaniv, Z., Dafni, A., & Palewitch, D. (1986). A preliminary classification of the healing potential of medicinal plants, based on a rational analysis of an ethnopharmacological field survey among Bedouins in the Negev Desert, Israel. Journal of Ethnopharmacology, 16(2–3), 275–287. https://doi.org/10.1016/0378-8741(86)90094-2
Gamboa, P. (2014). Arte precolombino, arte moderno y arte latinoamericano. Ensayos: Historia y Teoría del Arte.
Gobernación del Departamento del Valle del Cauca. (2021). Resultados anuales de la asistencia técnica del municipio de Caicedonia, Valle del Cauca. https://www.valledelcauca.gov.co/zar/publicaciones/70953/resultados-anual-de-la-asistencia-tecnica-del-municipio-de-caicedonia-valle-del-cauca/
Gómez, C., & Garrido, E. (2017). Iconografías y representaciones del jaguar en Colombia: De la permanencia simbólica a la conservación biológica. Antípoda. Revista de Antropología y Arqueología, 28, 131–152. http://www.scielo.org.co/scielo.php?pid=S1900-54072017000200131&script=sci_arttext
González, A. R. (1998). Arte precolombino. Cultura La Aguada, arqueología y diseños. Filmediciones Valero.
Gower, J. C. (1966). Some distance properties of latent root and vector methods used in multivariate analysis. Biometrika, 53(3-4), 325-338.
Guayasamín, J. M., Cisneros-Heredia, D. F., McDiarmid, R. W., Peña, P., & Hutter, C. R. (2020). Glassfrogs of Ecuador: Diversity, evolution, and conservation. Diversity, 12(6), 222. https://doi.org/10.3390/d12060222
Guayasamín, J. M., Frenkel, C., Varela-Jaramillo, A., & Ron, S. R. (2022). Centrolene buckleyi. En S.
R. Ron, A. Merino-Viteri, & D. A. Ortiz (Eds.), Anfibios del Ecuador (Versión 2024.0). Museo de Zoología, PUCE. https://www.bioweb.bio/faunaweb/amphibiaweb/FichaEspecie/Centrolene%20buckleyi
Guayasamín, J. M., Varela-Jaramillo, A., & Frenkel, C. (2023). Nymphargus griffithsi. En S. R. Ron,
A. Merino-Viteri, & D. A. Ortiz (Eds.), Anfibios del Ecuador (Versión 2024.0). Museo de Zoología, PUCE.
https://www.bioweb.bio/faunaweb/amphibiaweb/FichaEspecie/Nymphargus%20griffithsi Guevara-Molina, S. C., Benítez-Cubillos, E. L., & Londoño-Guarnizo, C. A. (2017). Colostethus
fraterdanieli (Silverstone, 1971). Rana silvadora. Catálogo de Anfibios y Reptiles de Colombia, 3(2), 14–19.
Heyer, M. M., Heyer, W. R., & de Sá, R. O. (2006). Leptodactylus fragilis (Brocchi): White-lipped thin-toed frog. Catalogue of American Amphibians and Reptiles, 830, 1.
Jolliffe, I. T., & Cadima, J. (2016). Principal component analysis: a review and recent developments. Philosophical transactions of the royal society A: Mathematical, Physical and Engineering Sciences, 374(2065), 20150202.
Legast, A. (1993). La fauna en el material precolombino Calima (Vol. 56). Fundación de Investigaciones Arqueológicas Nacionales.
Legast, A. (1998). La fauna muisca y sus símbolos. Boletín de Arqueología de la FIAN, 5–103. Legast, A. (2000). La figura serpentiforme en la iconografía muisca. Boletín Museo del Oro, 46, 22–39. Legendre, P., & Legendre, L. (2012). Numerical ecology (Vol. 24). Elsevier.
Lele, S., & Richtsmeier, J. T. (1991). Euclidean distance matrix analysis: A coordinate-free approach for comparing biological shapes using landmark data. American Journal of Physical Anthropology, 86(3), 415–427.
Márquez, C., Bechard, M., Gast, F., & Vanegas, V. H. (2005). Aves rapaces diurnas de Colombia.
Instituto de Investigación de Recursos Biológicos Alexander von Humboldt.
Morales, D. C. (1996). El simbolismo animal en la cultura medieval.
Moreno Arias, R. Á., Velasco, J. A., Urbina-Cardona, J. N., Cárdenas-Arévalo, G., Medina-Rangel, G. F., Gutiérrez-Cárdenas, P. D. A., … & Noguera-Urbano, E. (2023). Atlas de la biodiversidad de Colombia. Anolis.
Murtagh, F., & Contreras, P. (2012). Algorithms for hierarchical clustering: An overview. Wiley Interdisciplinary Reviews: Data Mining and Knowledge Discovery, 2(1), 86–97.
Navarro-Salcedo, P., Muñoz-Acevedo, S., & Bernal-Bautista, M. H. (2019). Pristimantis permixtus: Rana de muslos naranja. Asociación Colombiana de Herpetología – ACH. Catálogo de Anfibios y Reptiles de Colombia, 5(1), 40–46.
Ochante-Sauñe, M. (2015). Simbolismo de los sapos y culebras en la cosmovisión andina. Revista Ciberandes. http://www.ciberandes-magazin.com/2015/07/simbolismo-de-sapos-y-culebras-en-la-cosmovision-andina/
Ortiz, D. A., Coloma, L. A., Frenkel, C., Ron, S. R., Quiguango-Ubillús, A., & Páez-Rosales, N. (2022). Hyloscirtus larinopygion. En S. R. Ron, A. Merino-Viteri, & D. A. Ortiz (Eds.), Anfibios del Ecuador (Versión 2024.0). Museo de Zoología, PUCE. https://bioweb.bio/faunaweb/amphibiaweb/FichaEspecie/Hyloscirtus%20larinopygion
Paredes, C., Matos, H., de la Vega, R., & Pardo, E. (2019). Reconocimiento semiautomático de depresiones kársticas mediante análisis de agregados en sus distribuciones morfométricas. Aplicación al karst del Estado de Yucatán, México. Revista Mexicana de Ciencias Geológicas, 36(2), 270–288. https://www.redalyc.org/journal/572/57265251010/html/
Redi, Y. M. (2019). Ejercicio de caracterización zoológica de representaciones de felinos en piezas arqueológicas de la Colección Moreno (Museo de La Plata). Fragmentos del Pasado-do Passado, (6).
Ríos-Soto, J. A., Arango-Lozano, J., & Rivera-Molina, F. A. (2018). Micrurus mipartitus (Duméril, Bibron y Duméril, 1854): Rabo de ají, coral, coralillo, gargantilla, matagatos o mataganado. Catálogo de Anfibios y Reptiles de Colombia, 4(1), 37–44.
Ríos-Orjuela, J. C., Camacho-Bastidas, J. S., & Jerez, A. (2020). Appendicular morphology and locomotor performance of two morphotypes of continental anoles: Anolis heterodermus and Anolis tolimensis. Journal of Anatomy, 236(2), 252–273.
Shimodaira, H. (2002). An approximately unbiased test of phylogenetic tree selection. Systematic biology, 51(3), 492-508.
Shimodaira, H. (2004). Approximately unbiased tests of regions using multistep-multiscale bootstrap resampling. Ann. Statist. 32(6): 2616-264.
Shimodaira, H., & Hasegawa, M. (2001). CONSEL: for assessing the confidence of phylogenetic tree selection. Bioinformatics, 17(12), 1246-1247.
Silva de Lima, J., Azarias dos Santos, C. M., & Aquino dos Santos, C. K. (2020). Utilização da etnozoologia e educação ambiental para desvendar a concepção das crianças em relação aos anfíbios anuros [Use of ethnozoology and environmental education to unveil children’s conception of anuran amphibians]. Diversitas Journal, 5(2), 814–823. https://doi.org/10.17648/diversitas-journal-v5i2-726
Suzuki, R., & Shimodaira, H. (2006). Pvclust: An R package for assessing the uncertainty in hierarchical clustering. Bioinformatics, 22(12), 1540–1542. https://doi.org/10.1093/bioinformatics/btl117
Tocarema, G. (2018). Molá; Toponimia de los sitios sagrados del pueblo pijao en el municipio de Natagaima del Departamento del Tolima [Trabajo de grado en Antropología]. Universidad Externado de Colombia.
Uribe-Villegas, M. A. (1991). La orfebrería Quimbaya tardía: Una investigación en la colección del Museo del Oro.
Uribe-Villegas, M. A. (2016). Las aves en la orfebrería prehispánica del Cauca Medio.
Vargas-Salinas, F., Rodríguez-Collazos, L. A., & Suárez-Mayorga, Á. M. (2014). Andinobates bombetes (Myers y Daly, 1980) Rana rubí o rana ají. Catálogo de Anfibios y Reptiles de Colombia, 2(2), 13–18.
Vargas-Salinas, F., Atehortua-Vallejo, M. A., Arcila-Pérez, L. F., Jiménez-Vargas, G. M., González-Acosta, C., Casas-Cardona, S., & Grajales-Echeverry, A. (2020). Andinobates bombetes.
Ward, J. H. (1963). Hierarchical grouping to optimize an objective function. Journal of the American Statistical Association, 58(301), 236–244. https://doi.org/10.1080/01621459.1963.10500845
Williams, K. L. (1994). Lampropeltis triangulum. Catalogue of American Amphibians and Reptiles, 594, 1–5.
Yánez-Muñoz, M. H., Reyes-Puig, J. P., Batallas-Revelo, D., Broaddus, C., Urgilés-Merchán, M., Cisneros-Heredia, D. F., & Guayasamín, J. M. (2021). A new Andean treefrog (Amphibia: Hyloscirtus bogotensis group) from Ecuador: An example of community involvement for conservation. https://d1wqtxts1xzle7.cloudfront.net/81981122/11914-libre.pdf

Comentarios recientes